Steven Smith, Ph.D.

- Genome Evolution
- Role of Non-B Structures in Epigenetics, Epimutation and DNA Damage
Role of Non-B Structures in Epigenetics, Epimutation and DNA Damage
The working hypothesis in my laboratory is that genomes, like other components of living things are the subject to evolution by natural selection. This line of investigation is a tradition at City of Hope that was founded by Susumu Ohno.
My laboratory has explored the genetic and epigenetic aspects of genome evolution linked to a class of DNA sequences that are dynamic in the sense that in addition to normal Watson-Crick B-DNA, they can form G-quadruplex, i-motif and triplex DNA structures (Figure 1). Considerable evidence now shows that these dynamic sequences perform epigenetic functions that are important in gene regulation at the level of transcription and translation. However, adoption of these sequences in epigenetic roles comes with a cost, since their adventitious formation during DNA replication can lead to serious mutations, deletions and rearrangements.
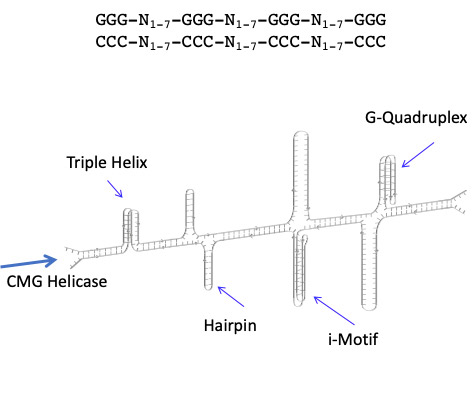
Figure 1: Structures associated with the sequence GGG-N1-7-GGG-N1-7-GGG-N1-7-GGG. Replication through these sequences is preceded by CMG helicase action.
The advent of sequences capable of formation of these canonical structures and the intermediates that form them in DNA genomes appears to have taken place rather late in biological evolution, with biota from the Ediacaran epoch [635-541 million years before present] likely to be the first eukaryotic organisms capable of tolerating them. That the increasing frequency of these sequences tracks the evolution of biological complexity in the animal kingdom strongly supports an epigenetic role associated with them Figure 2.
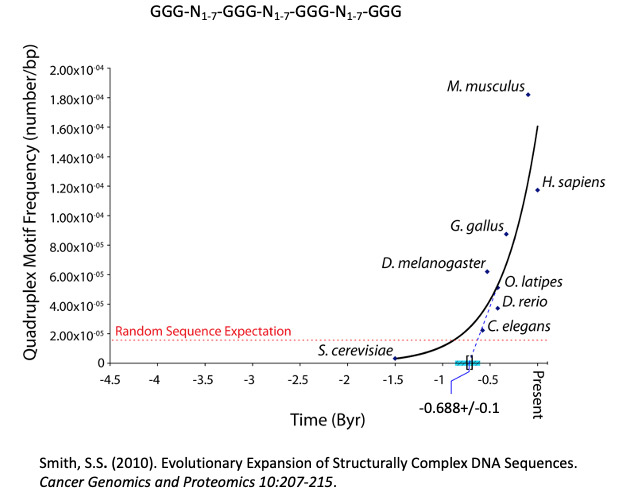
Figure 2: The frequency of GGG-N1-7-GGG-N1-7-GGG-N1-7-GGG Sequences Increases with Organism Complexity in the Animal Kingdom. This increase again suggests that the sequences are under positive selection and may play a role in epigenetics. Extrapolation of the graph to the time axis suggest an origin near the beginning of the Edicaran Epoch. On the other hand, among the most complex organisms, loss of control of the formation of these sequences can result in spontaneous mutagenesis. For example, in recent work we showed that one class of deletions was associated with a wide spread repetitive element of the form (TCCC)n. This element spontaneously forms an i-motif under physiological conditions when n is greater than 8, and we noted deletions at RACK7 (n=9), ABL1 (n=11) and PLA2GA (n=15) in human cancer cells (Figure 3).
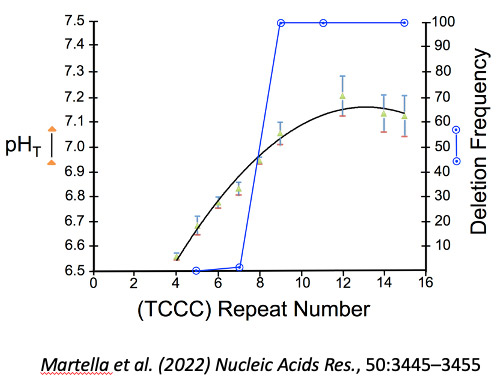
Figure 3: Length dependence of i-motif formation at neutral pH correlated with deletion frequency. pHT value: closed triangles (green) +/- standard deviation determined from the titration curve fit. Fraction of cloned sequences with a deletion: open circles (blue).
When we looked at DNA from bone marrow donors and prostate DNA specimens from patients who did not have cancer, we noted that the deletions at RACK7 appeared to become more frequent and more extensive with in older patients (Figure 4).

Figure 4: RACK7 sequences obtained from patients without a diagnosis of cancer. (A) DNA from the bone marrow of a healthy 22-year-old bone marrow donor. (B) DNA from peripheral blood B-Cells from a healthy 41-year-old bone marrow donor. (C) DNA from peripheral blood B-cells from a healthy 60-year-old bone marrow donor. (D) DNA from a 69-year-old prostate patient with a negative biopsy for prostate cancer with high-grade prostatic intraepithelial neoplasia (HGPIN). (E) DNA from a 74-year-old prostate patient with a negative biopsy for prostate cancer with proliferative inflammatory atrophy (PIA). Sequences from the GCSkew region of each cloned representative are shown for clarity. Regions flanking the region of GC-skew match the reference sequence () given in GRCh38.p12. However, in the region of the concatemer [(CCTG)8-CC-(TCCC)9-(TTCC)9], none of the sequences from these patients matches the reference sequence () for RACK7 from Human chromosome 20, GRCh38.p12 primary assembly.
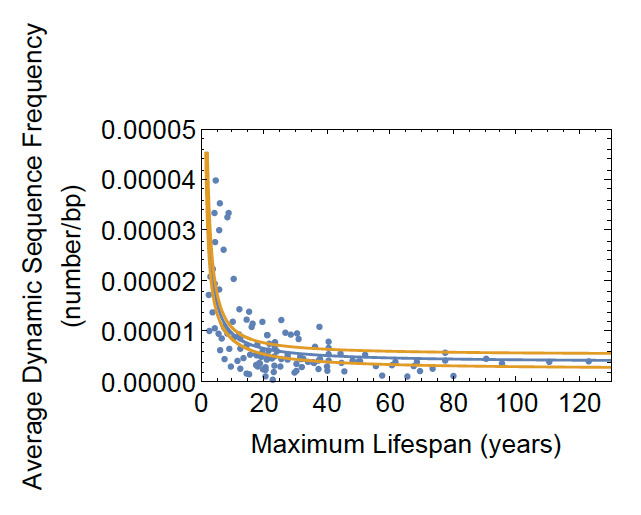
This in turn suggests that the genomic frequencies of dynamic sequences may set spontaneous mutation rates associated with aging. As a test of that idea we studies frequencies of several mutagenic dynamic sequences as a function of mammalian lifespan. Consistent with that idea the frequencies decrease dramatically with increased lifespan among the mammals (Figure 5).
Smith, S.S. (2025) The Influence of CG sites on Dynamic DNA Sequence Mutagenesis in the Genomic Evolution of Mammalian Lifespan. Nucleic Acids Research 53 , gkaf762 https://doi.org/10.1093/nar/gkaf762.
Figure 5: Correlation between the Average Frequency of a Set of Six Dynamic Sequences and Lifespan for 126 representative mammalian genomes. The evolutionary rise of dynamic sequences (Figure 2) overall suggested that all such sequences cannot be under negative selection, and our most recent work shows that there is a second class of dynamic sequences that contain one or more CG sites that is under positive selective pressure with lifespan in the mammals (Figure 5). This suggests that the spontaneous mutagenicity of these sequences is suppressed by DNA methylation and methylated DNA binding proteins, permitting them to perform needed epigenetic functions, and leading to a model for age linked mutagenesis (Figure 6).
Smith, S.S. (2025) The Influence of CG sites on Dynamic DNA Sequence Mutagenesis in the Genomic Evolution of Mammalian Lifespan. Nucleic Acids Research 53 , gkaf762 https://doi.org/10.1093/nar/gkaf762.
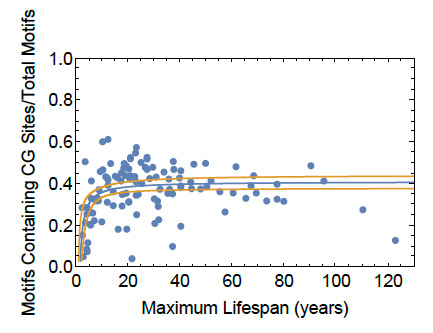
Figure 6: Fraction of G3-4N1−7G3-4N1−7G3-4N1−7G3-4 Dynamic Motifs Containing One or More CG Sites. Data for the 126 representative mammalian genomes studied shows an upward trend in the fraction of motifs containing at least one CG site with lifespan.
Smith, S.S. (2025) The Influence of CG sites on Dynamic DNA Sequence Mutagenesis in the Genomic Evolution of Mammalian Lifespan. Nucleic Acids Research 53 , gkaf762 https://doi.org/10.1093/nar/gkaf762
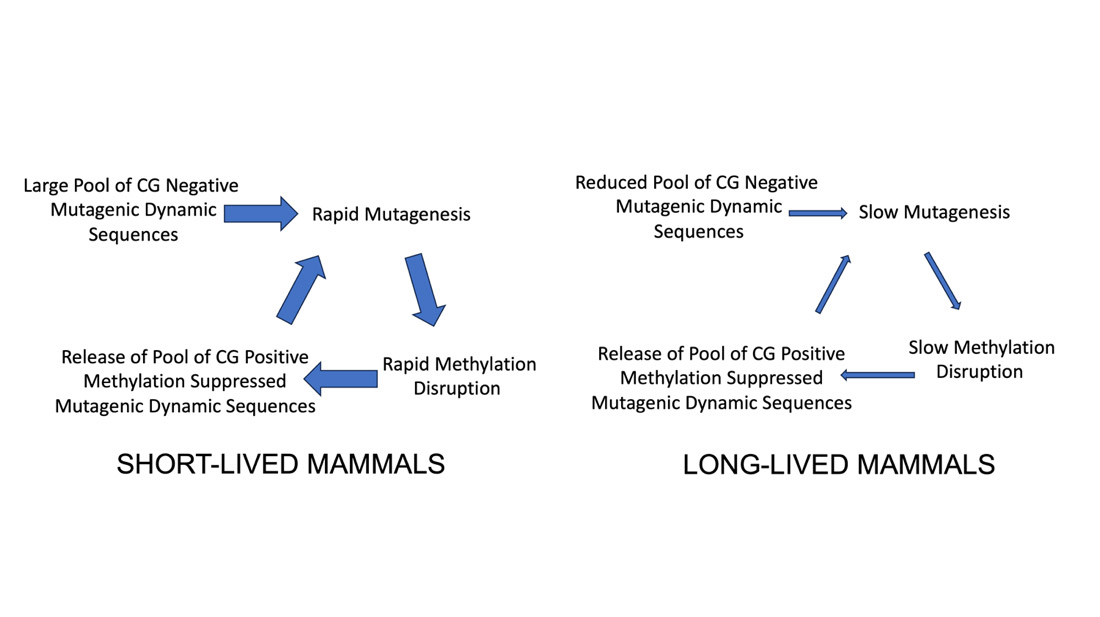
Figure 7. Contribution of Mutagenic Dynamic Sequences to the Species-Specific Mutation Rate and the Species-Specific Methylation Rate. In addition to spontaneous deamination, and replication errors, dynamic sequences are expected to contribute to both endogenous mutation accumulation and methylation pattern disruption based on both differences in genomic frequencies in the CG positive and CG negative pools. In short lived mammals, mutagenesis initiated from the large CG negative pools would rapidly disrupt methylation and structure suppression in the CG positive pool releasing that pool for additional mutagenesis. Thus methylation disruption would continue until both pools are depleted. In long-lived mammals with small CG negative pools, the release of CG positive sequences from methylation suppression would play out more slowly over a longer lifespan. In both long-lived and short-lived mammals, methylation suppression of structure formation will cause promoter and exon sequences to experience lower rates of dynamic structure linked mutagenesis. However, methylation will not inhibit transcription associated mutagenesis by mutagenic dynamic sequences.
Current research is aimed at determining whether the newly discovered function of DNA methylation in the suppression of Dynamic Sequence mutagenesis, preceded its role in gene expression.
Professor Smith’s publications can be viewed at Research gate.
Biomedical Research Center
1500 East Duarte Road
Duarte, CA 91010
Location
Duarte Cancer Center
Duarte, CA 91010
Education & Experience
Degrees
- 1974, University of California, Los Angeles, CA, Ph.D., Molecular Biology
- 1968, University of Idaho, Moscow, ID, B.S., Zoology
Professional Experience
- 2022 - present. Professor Emeritus, Department of Stem Cell Biology and Regenerative Medicine, City of Hope, Monrovia, CA
- 2016 - 2022 Professor Emeritus, Department of Department of Hematologic Malignancies Translational Science, City of Hope, Monrovia, CA
- 2001 - 2016, Professor of Molecular Science, Division of Urology/Department of Surgery, City of Hope, Duarte, CA
- 1990 - 2001, Director, Department of Cell & Tumor Biology, Division of Surgery, City of Hope, Duarte, CA
- 1990 - 1998 Director, Program in Molecular Carcinogenesis, NCI-designated Clinical Cancer Center, City of Hope National Medical Center, Duarte, CA
- 1995-2000 Research Scientist, Division of Surgery, City of Hope National Medical Center, Duarte, CA
- 1987 - 1995, Associate Research Scientist, Division of Surgery, City of Hope, Duarte, CA
- 1985 - 1987, Assistant Research Scientist, Department of Thoracic Surgery, Division of Surgery, City of Hope, Duarte CA
- 1982 - 1984, Assistant Research Scientist, Department of Molecular Biology, City of Hope, Duarte, CA